 The retino-cortical mapping
The retino-cortical mapping
RESEARCH
The main focus of our research is to use mathematical modeling and analysis to understand how the brain functions as a complex dynamical system. This work involves many different levels of description from the molecular basis of memory and learning at an individual synapse, to the physiological mechanisms underlying neuronal firing, to the properties of local circuits processing elements of a visual scene for example, to the large-scale structure of cortex responsible for higher cognitive function.
| The retino-cortical mapping
| |||
Superimposed upon this retinotopic map are a number of additional (approximately) periodic feature maps reflecting the fact that neurons respond preferentially to various aspects of a stimulus such as its orientation and left/right eye (ocular) dominance. Orientation preference changes continuously as a function of cortical location except at singularities or pinwheels that tend to be located at the center of ocular dominance columns. Around each orientation pinwheel is a ring of orientation selective cells.
 LEFT: Distribution of orientation patches revealed by
optical imaging [Courtesy of Gary Blasdel] RIGHT: Orientation
pinwheels LEFT: Distribution of orientation patches revealed by
optical imaging [Courtesy of Gary Blasdel] RIGHT: Orientation
pinwheels
| 
| ||
The laterally spreading connectional fields made by pyramidal cells in superficial layers of V1 are broken into regularly spaced patches. These patchy lateral connections, which can extend up to 6mm in cortex, appear to link neurons with common functional properties such as orientation preference. Ongoing work by Dr. Alessandra Angelucci and Dr. Jennifer Lund at the Moran Eye Center, University of Utah, has revealed that feedback connections from higher cortical areas such as V2, V3 and MT appear to obey similar rules.
 The patchy nature of lateral and feedback long-range
connections in primate V1. [Courtesy of Dr. Alessandra Angelucci and Dr.
Jennifer Lund (University of Utah)] The patchy nature of lateral and feedback long-range
connections in primate V1. [Courtesy of Dr. Alessandra Angelucci and Dr.
Jennifer Lund (University of Utah)]
| |||
The long-range lateral and feedback connections are thought to provide a substrate for center-surround interactions. The surround fields are regions beyond the classical receptive field of a neuron where stimuli do not drive the cell, but modulate its response to stimuli placed within its classical receptive field. One common experimental paradigm is to investigate the response to stimuli consisting of circular (center) and annular (surround) gratings of differing contrasts, orientations and diameters. When both center and surround are stimulated with high contrast gratings close to the orientation of the unit's peak tuning response, the surround typically suppresses the center unit's tuning response. However, the degree of suppression tends to decrease, and sometimes becomes facilitatory, for surround stimulations at orientations sufficiently dissimilar to the preferred orientation of the center or at low contrasts.

| Center-surround modulation | ||
We are currently analyzing a large-scale dynamical model of primary visual cortex in order to understand the role of long-range lateral and feedback connections in modulating both the classical receptive field, which expands at low contrasts, and the inhibitory surround. [In collaboration with Dr. Jack Cowan, University of Chicago]. These processes are extremely important, since they provide a substrate for V1 to integrate spatial information across significant parts of the visual field, thus contributing to the creation of a global visual percept from local features. Such integration has traditionally been attributed to higher visual areas that are known to have larger classical receptive fields.
 LEFT: Spiral hallucination RIGHT: Spiral
hallucination LEFT: Spiral hallucination RIGHT: Spiral
hallucination
| 
| ||
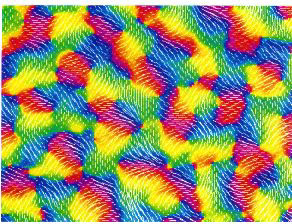
Our work has focused on a continuum model of V1 in which cells signal both the position and orientation of a local stimulus. Using symmetric bifurcation theory, we have shown that the cortical activity patterns underlying common visual hallucinations can be accounted for in terms of certain symmetry properties of the lateral connections, specifically, that they are invariant under the action of the planar Euclidean group - the group of rigid motions in the plane - rotations, reflections and translations. The resulting representation is twisted due to an anisotropy in the lateral connections, which tends to favor directions that are correlated with the orientation preferences of the interacting cells. [In collaboration with Dr. Jack Cowan (University of Chicago) and Dr. Martin Golubitsky (University of Houston)]
 LEFT: Pressure phosphene RIGHT: Honeycomb
hallucination LEFT: Pressure phosphene RIGHT: Honeycomb
hallucination
| 
| ||
In order to account for more complex hallucinations, we will need to extend our theory to incorporate additional properties of images such as color, depth and motion perception.
 LEFT: Lattice tunnel hallucination RIGHT: Tunnel
hallucination LEFT: Lattice tunnel hallucination RIGHT: Tunnel
hallucination
| 
| ||
| |||
The dependence of wave velocity on the underlying pattern of synaptic connectivity has been analysed in the case of homogeneous networks. However, certain care must be taken in interpreting these results since traveling wave solutions are sensitive to the degree of homogeneity in the connectivity pattern, particularly in the case of slowly moving waves. This is a consequence of the fact that traveling wavefronts are not structurally stable solutions, that is, they correspond to heteroclinic orbits of an equivalent dynamical system. We have recently used homegenization theory to show that in a simplified one-dimensional cortical model, inhomogeneities induced by long-range lateral connections can lead to propagation failure. Moreover, the conditions for propagation failure are sensitive to the precise structure of the weight distribution.

| Arnold tongues for a sinusoidally forced integrate-and-fire neuron | ||
For more information contact Paul C. Bressloff, 5-1633